Enlaces directos de pirofosfato del ATP en la glucólisis
Enviado por Chuletator online y clasificado en Medicina y Salud
Escrito el en  español con un tamaño de 49,01 KB
español con un tamaño de 49,01 KB
ETAPA 1 - Glucogenolisis -
glucosa a piruvatoETAPA 2 - Complejo Piruvato deshidrogenasa-
Piruvato a Acetil-CoAETAPA 3 - Ciclo de Krebs - Acetil CoA a CO2 y H2O a ATP
ETAPA 4 - cadena transportadora de electrones
Glucólisis
Degradación, ruptura, lisis, oxidación de la glucosa. Primera parte del proceso catabólico.
De una glucosa se puede obtener una molécula degradada llamada piruvato.
Luis Pasteur: comprobó que se podía obtener el CO2 y etanol pero no sabía con exactitud de donde
Hermanos Buchner: comprobaron que se podía obtener un CO2 y etanol de una molécula de sacarosa en presencia de levaduras
Embed, Meyerhaaf, Parnas y Warburg: descifraron que en efecto el CO2 y etanol se podían obtener a partir de los azúcares o de la sacarosa (glucosa) a partir de 10 reacción con una enzima cada una, llamado Glucólisis o Vía de Embed, Meyerhaaf, Parnas y WW.
La glucólisis se puede dar en todos los tejidos, ya que se puede dar en ambientes anaeróbicos y aeróbicos. Lo que significa que al degradar la glucosa, tenemos tres vías:
Vía aeróbica: que implica que la glucosa puede formar CO2, H2O y ATP
Vía anaeróbica: implica la ausencia de oxígeno, en los músculos, en los eritrocitos, formando lactato
Vía anaerobica que se identifica en las bacterias: produce etanol
La glucólisis es una fuente de energía potencial porque los enlaces que contienen la glucosa, son enlaces que pueden liberar energía. Cuando la glucosa se rompe se va liberando energía para el cuerpo.
La glucólisis y sus 10 reacciones está dividida en dos etapas
Etapa preparatoria - reacción 1-5:
4 mecanismos importantes
Gasto de 2 ATP: se usa la energía que se libera al romper el grupo fosfato del ATP, y vamos a usar ese grupo fosfato liberado para que sirva de manera estructural
Preparar glucosa para que sea oxidada: adicionando grupos fosfato
Se obtiene gliceraldehido y dihidroxiacetona
Se obtienen 2 moléculas de gliceraldehido-3-fosfato
Etapa beneficiosa - reacción 6-10 -
ganancia de energía de dos tipos:
Energía en forma de electrones: en un proceso oxidativo
Energía en forma de nucleótidos de alta energía - ATP
ganancia de 2 moléculas de piruvato
IMPORTANCIA BIOMÉDICA
Piruvato cinasa: enzima que se aumenta drásticamente cuando hay anemia hemolítica, cuando hay destrucción de eritrocitos porque los propios anticuerpos los están atacando
Fosfofructoquinasa: regula la glucólisis de manera alostérica, se ve alterada en procesos de falta de oxigenación cerebral (mareos, ganas de vomitar)
Aldolasa: se asocia a nivel muscular, se altera drásticamente cuando hay problemas neuroendogenos y debilidad muscular y la enfermedad de Guillain-Barre (desmineralización de las neuronas y no llevan el conducto nervioso, provocado por un virus)
REACCIONES ETAPA PREPARATORIA
Glucosa ya está en la célula, por medio de glut, pero para evitar que se quede ahí y no salga, el cuerpo marca a esa glucosa
FOSFORILACIÓN de la glucosa, para que no pueda salir de la célula, con un fosfato libre que viene de un ATP que rompíó sus enlaces por medio de una transferasa llamada hexocinasa o hexoquinasa con su cofactor magnesio. La hexocinasa transfiere este grupo fosfato libre a la posición 6 de la glucosa, creando la uníón del grupo fosfato, creando la glucosa-6-fosfato
HEXOCINASA: libre en el citoplasma - en todos los tejidos - enzima alostérica - fosforila hexosas incluyendo a la glucosa - reacción irreversible - ΔG negativo - libera alta cantidad de energía - primera enzima reguladora de la glucólisis - baja Km - alta afinidad enzima sustrato - baja velocidad máxima - no fosforila moléculas en alta cantidad sino solo las necesarias - cuando hay baja cantidad de glucosas las fosforila fácilmente - Si la concentración de glucosa-6-fosfato o de ATP o de Acetil CoA (molécula que permite que la glucosa entre al ciclo de Krebs) es alta, la hexocinasa se inhibe sólita (es autorregulable, regula la entrada de la glucosa a la célula), cuando la concentración es baja se vuelve a activar para continuar con la reacción
GLUCOCINASA: almacenada en el núcleo por una proteína llamada proteína receptora de glucosinasas que hace que salga hacia el hígado, páncreas e hipotálamo cuando hay alta cantidad de glucosa - fosforila solo glucosa - alta Km - alta Velocidad máxima - menor afinidad enzima sustrato - cuando hay alta cantidad glucosas las fosforila fácilmente y fosforila cuantas glucosas encuentre - cuando ya no haya alta cantidad de glucosa, se vuelve a almacenar por la proteína receptora de glucosinasas en el núcleo - se inhibe por la baja concentración de glucosa en el citoplasma o por la alta presencia de fructosa-6-fosfato y se vuelve a activar por la alta concentración de glucosa y baja presencia de fructosa-6-fosfato
ISOMERIZACIÓN: tenemos la glucosa-6-fosfato y necesitamos que se isomeriza en una fructosa-6-fosfato, por medio de la glucosa-6-fosfato-isomerasa o fosfohexosa isomerasa que posee 3 aminoácidos en su estructura (Lys(rompe anillo de la glucosa), glu (reorganiza los enlaces), his (cierra anillo de la fructosa)) para generar el proceso de isomerización (no son coenzimas) - libera poca energía - ΔG+ - reacción reversible - no tiene reguladores - enzima isosterica
SEGUNDA FOSFORILACIÓN: utilizamos otro ATP, se rompe el enlace liberando mucha cantidad de energía y se transfiere el grupo fosfato que se líberó hacia la posición 1 de la fructosa por medio de una fosfofructocinasa 1 con la ayuda del magnesio formando fructosa-1,6-bifosfato. Se libera mucha energía - ΔG negativo - reacción irreversible - enzima alostérica - segunda enzima reguladora de la glucólisis (regula la velocidad de la glucólisis) - es regulada porque cuando hay aumento de ATP o de citrato (ciclo de krebs) se inhibe - se activa por la alta concentración de AMP y por la baja concentración de citrato - también se puede regular por glucogenólisis, mucha glucosa puede regular.
La enzima ALDOLASA rompe el anillo de la fructosa 1,6-difosfato en dos, produciendo un gliceraldehido 3 fosfato (listo para oxidarse pasa a la reacción 6) y una dihidroxiacetona fosfato (no está lista para oxidarse)
Aldolasa: enzima isosterica, no regulable - ΔG + - reacción reversible. Existen dos tipos
Aldolasa A: en músculos, específica para fructosa-1,6- bifosfato, es la de mayor proporción, genera el gliceraldehido-3-fosfato y la dihidroxiacetona fosfato
Aldolasa B: en hígado, puede ser utilizada por la fructosa-1,6-bifosfato y fructosa-6-fosfato
La dihidroxiacetona fosfato necesita isomerizar interconvertirse para poder convertirse en una aldosa para poder oxidarse, por medio de la enzima triosa fosfato isomerasa: enzima isosterica, no regulable - ΔG+ - reacción reversible - aun así la reacción tiene más afinidad por el sustrato que por el producto, entonces busca quedarse más como cetosa que como aldosa.
RESUMEN ETAPA PREPARATORIO
Entra en la reacción una glucosa, salen 2 moléculas de gliceraldehido 3 fosfato, ingresaron 2 moléculas de ATP y salen dos moléculas de ADP, hay gasto energético
REACCIONES ETAPA BENEFICIOSA
Ya tenemos dos moléculas de gliceraldehido 3 fosfato
Oxidación: del gliceraldehido 3 fosfato por medio de la enzima Gliceraldehido 3-fosfato-deshidrogenasa, esta quita hidrógenos. Es una enzima tetramérica, ya que tiene 4 monómeros conformados por 3 elementos importantes
NAD+: se reduce al captar el hidrógeno que la cisteína eliminar del gliceraldehido 3-fosfato
Cisteína: va a favorecer a la oxidación, eliminando o rompiendo el enlace del hidrógeno del gliceraldehido-3-fosfato (este se oxida)
Histidina: regulara el pH
Se sustituye el hidrógeno que se eliminó por un grupo fosfato inorgánico del citoplasma en la posición 1 del gliceraldehído, produciendo un 1,3 difosfoglicerato
En resumen: se oxida el gliceraldehido (el NAD se reduce), se intercambia electrones en forma de hidrógenos por un fosfato inorgánico
Cuando se satura el NAD, la estructura del gliceraldehido 3 fosfato deshidrogenas se relaja (configuració R), por lo que el NAD se va a la cadena transportadora de electrones, se separa del gliceraldehido 3 fosfato deshidrogenasa y este se tensa (configuración Tau), llega un nuevo NAD+ y está lista para volver a ser oxidada y así va acumulando electrones para la cadena transportadora de electrones.
Esta es una reacción reversible porque el NAD está en constante movimiento - ΔG+ - enzima isosterica - la pueden inhibir competitivamente 4 elementos: arsénico, flúor (se une en la posición (1) del grupo fosfato, ataca al producto, deteniendo la reacción y ya no pueda continuar en el proceso glucolítico de las bacterias y estas mueran, por esto se utiliza el flúor en las pastas dentales), Mercurio y cobre.
Cuando se une el grupo fosfato al gliceraldehído 3 fosfato se forma un enlace de alta energía, este se rompe y sirve para que este grupo fosfato del ahora 1,3 difosfoglicerato se una a un ADP que este sólito y forme un ATP, por medio de una fosfoglicerato cinasa con ayuda de magnesio. Este proceso se llama fosforilación a nivel de sustrato. Produciendo un 3 fosfoglicerato
Esta es una reacción reversible - ΔG + - la fosfoglicerato cinasa es la única enzima cinasa que es reversible, porque los dos enlaces están aportando la misma cantidad de energía, entonces se puede tomar un grupo fosfato de un ADP para regenerar la molécula de 1,3 difosfoglicerato
A nivel de los ERITROCITOS en la glucólisis no solo se dan 10 reacciones, sino que 11, se relaciona con el proceso respiratorio. En el eritrocito tenemos un 1,3 difosfoglicerato y una enzima bisfosfoglicerato mutasa que traslada el grupo fosfato de la posición 1 del 1,3 difosfoglicerato a la posición 2, formando un 2,3 difosfoglicerato, cuya función es que la hemoglobina pierde afinidad por el oxígeno. Esta es una reacción momentánea. Luego existe otra enzima llamada 2,3 bisfosfoglicerato fosfatasa que romperá el grupo fosfato de la posición 2 de la 2,3 difosfoglicerato para obtener un 3 fosfoglicerato.
PREGUNTA: ¿Qué relación tiene la glucólisis con la respiración? Cuando comemos hay proceso glucolítico, se forma un 2,3 difosfoglicerato favoreciendo la liberación de oxígeno de la hemoglobina
ISOMERIZACIÓN: cambiar el grupo fosfato de la posición 3 del 3 fosfoglicerato a la posición 2, isomerización con el fin de formar otro enlace de alta energía, por medio de la fosfoglicerato mutasa con ayuda de magnesio, formando un 2 fosfoglicerato
Es una reacción reversible - ΔG+ - enzima alostérica
DESHIDRATACIÓN: se empieza a formar el enlace de alta energía, por medio de la enzima enolasa. Romperá el enlace del H + y OH- para liberar un H2O, generando un alqueno con un enlace de alta energía llamado fosfoenolpiruvato.
Reacción reversible - lo inhibe el flúor o PO4 formando una molécula de flúor-fosfato-magnesio que genera un proceso competitivo, deteniendo la reacción (si no hay magnesio, el flúor no se puede unir al grupo fosfato
El FLÚOR inhibe la acción de la enolasa y la reacción ya no funciona y no haya proceso glucolítico en la bacteria y esta muere
FORMACIÓN DE PIRUVATO: transferir el grupo fosfato con enlace de alta energía del fosfoenolpiruvato a un ADP, por medio de una piruvato cinasa, formando como producto piruvato y ATP
El piruvato genera tautómeros: moléculas que pueden existir en un mismo momento como dos moléculas distintas. Isómeros que tienen dos formas estereoquímicas en un mismo momento ya que se forma una cetona o un alcohol (ceto-enol)
Reacción irreversible - isostérica
REGULACIÓN DE LA Glucólisis
ENZIMAS REGULADORAS ISOSTÉRICAS
Hexocinasa (reacción 1 - formación de glucosa 6 fosfato)
alosterismo: regulación con respecto a la concentración de sustrato o producto
homotrópico: de la misma vía
heterotópico: molécula de otra vía regula la vía metabólica
La glucosa que estamos usando viene de afuera de la célula no del proceso como tal, que va a facilitar el mecanismo de la hexocinasa cuando hay alta concentración, entonces es regulación alostérica heterotópica
La hexocinasa regula la glucólisis porque identifica la concentración de glucosa
Puede regular otras vías
Fosfofructocinasa 1 (reacción 3 - formación de fructosa 1,6-difosfato)
Esta pertenece a un complejo enzimático, pertenece a un grupo donde hay más de una enzima unidas en la misma estructura, entre las cuales está la fosfatasa, al complejo se le llama PFK-2 fosfofructocinasa 2
La fosfofructocinasa 1 funciona por modificación covalente: adición o eliminación de un fosfato (fosforilación) al sitio activo de la molécula para activar o inhibir la enzima.
INSULINA (hormona): cuando aumenta la concentración de glucosa a nivel sanguíneo (consumo de alimentos) hay una secreción de insulina, que es un mensajero primario que cuando la glucosa llega a los receptores celulares se sintetiza dentro de la célula una fructosa-2-6-bifosfato (mediadora), que al haber mucha concentración de ella, indicará a la fosfofructocinasa 2 que se desfosforile, perdiendo sus grupos fosfatos y activándose la fosfofructocinasa 1 permitiendo que se lleve a cabo la glucólisis. La fosfatasa del complejo PFK-2 se fosforila también y se inhibe. La función de la fosfatasa es sintetizar una nueva molécula de glucosa (gluconeogénesis)
Cuando la glucosa ya está disminuyendo y se forma glucagón (hormona). Entonces la fructosa-2,6-bifosfato disminuye y ya no desfosforila a la PFK-2 sino que se fosforila, inhibiendo a la fosfofructocinasa 1 deteniendo la glucólisis y activando la fosfatasa sintetizando nuevas moléculas de glucosa en la gluconeogénesis.
Piruvato cinasa (reacción 10 - formación de piruvato): regulada por glucagón, cuando hay alta concentración de glucagón la enzima se inhibe, cuando hay baja concentración de glucagón la enzima se activa. Cuando el glucagón se une al receptor de membrana celular (proteína G) se activa la enzima de la proteína G llamada adenilciclasa que rompe dos dos enlaces de alta energía del ATP para formar un AMP como molécula de baja energía, se vuelve un segundo mensajero para tener un AMPc que activa a la enzima proteincinasa que permitirá que un grupo fosfato se una a la piruvato cinasa, inhibiéndola e inhibiendo la glucólisis
También la puede regular la alanina, porque ella puede llegar a formar glucosa en el hígado, entonces una alta o baja puede favorecer la inducción o represión de la piruvato cinasa.
También lo regula el calcio o los ácidos grasos o el ATP, la alta concentración de cualquiera de ellos puede inhibir a la piruvato cinasa.
La alta concentración de fosfoenolpiruvato y la fructosa 1,6-bifosfato la pueden activar.
La piruvato cinasa necesita a los cofactores potasio y manganeso para poder realizar la reacción
Si la piruvato cinasa se disminuyera se identificará un problema en los eritrocitos, ya que disminuye su glucólisis y no haya síntesis de 2,3 difosfoglicerato, provocando que no haya liberación de oxígeno, este se quedara atrapado en la hemoglobina y este no podrá salir a los tejidos que debe llegar, provocando falta de aire o anemia.
La piruvato cinasa, la glucosa-6- fosfato isomerasa y la glucosa 6 fosfato deshidrogenasa se ven alteradas en la anemia, almacenando oxígeno en la hemoglobina
DESTINOS DEL PIRUVATO
Destino aeróbico
Destino anaeróbico bacterias(formación de etanol - fermentación)
Tenemos piruvato de 3 carbonos y el etanol es de 2 carbonos por lo que para formar este etanol el piruvato tiene que perder un carbono (descarboxilación) en forma de CO2, por medio de la piruvato descarboxilasa, pero aquí forma un acetaldehído (tóxico) este se reduce cuando un NADH+H dona electrones al aldehído (por un alcohol deshidrogenasa) para ahora sí formar etanol
Destino anaeróbico músculo - formación de ácido láctico o lactato - los músculos están acostumbrados a las actividades normales que se hace y de acuerdo a eso tiene cierta cantidad de oxígeno, pero cuando hacemos actividades fuera de lo normal, este oxígeno disminuye, creando un ambiente anaeróbico, por lo que no se puede hacer ciclo de krebs ni fosforilación oxidación aumentando el NADH y se empieza a acumular. Entonces la lactato deshidrogenasa traslada el hidrógeno del NADH al piruvato para obtener NAD + y poder seguir haciendo glucólisis. Pero cuando el piruvato recibe estos H se convierte en lactato.
Cuando uno se sigue moviendo el ácido láctico se dirige hacia el hígado por la vía sanguínea, aquí el ácido láctico devuelve a los H al NAD+ y regresa a ser piruvato, este piruvato en el hígado genera gluconeogénesis, formando glucosa que tomara la vía sanguínea y llegando al músculo.
El lactato hace que disminuya el pH (ácido) y por eso duele el músculo.
Si se sigue realizando esta actividad como ejercicio la mitocondria aumenta de tamaño y se gana resistencia muscular ya que se tendrá mayor capacidad de captación de oxígeno.
La lactato deshidrogenasa aumento en el corazón sirve para identificar un infarto agudo en el miocardio.
Puede formar un émbolo y para identificar problemas de coagulación.
DESCARBOXILACIÓN OXIDATIVA - COMPLEJO PIRUVATO DESHIDROGENASA
5 REACCIONES
El que va a empezar esta reacción es un complejo multienzimático (conjunto de enzimas interrelacionadas no covalentemente) llamado complejo piruvato deshidrogenasa, sus mecanismos de acción son ordenados, enzima 1 hace su efecto y la 2 y 3 están detenidas
Los complejos multienzimáticos tienen 3 carácterísticas importantes
Disminuyen las reacciones colaterales: permiten que la reacción sea ordenada, no todas al mismo tiempo.
Es un mecanismo coordinado
No permite colisiones externas
En el cuerpo existen 3 complejos enzimáticos importantes
Complejo piruvato deshidrogenasa: es el más importante porque se asocia a la oxidación de carbohidratos
Complejo alfa cetoglutarato deshidrogenasa: en el ciclo de krebs
Complejo alfa cetoácido deshidrogenasa: en la oxidación de aminoácidos
IMPORTANCIA BIOLÓGICA DEL COMPLEJO PIRUVATO DESHIDROGENASA
Carácterísticas
Tenemos un piruvato de 3 carbonos y el producto final será un Acetil-coA de 2 carbonos, que entrara en el ciclo de Krebs
Liberación de 1 CO2
Obtener energía en forma de electrones por lo que necesitamos un NAD+ que capte electrones y se convierta en NADH que llevará los electrones a la cadena transportadora de electrones y por fosforilación oxidativa generar un ATP
QUE ES EL COMPLEJO PIRUVATO DESHIDROGENASA - 5 REACCIONES
Este complejo contiene 5 enzimas de las cuales:
3 son ejecutoras: son las encargadas de generar la reacción en la que obtenemos energía en forma de electrones, obtenemos molécula de Acetil-CoA y molécula de CO2. Favorecen o reacción redox o reacción de transferencia (oxidoreductasas o transferasas), con ayuda de coenzimas (moléculas orgánicas ayudadoras) que facilitarán la transferencia de grupos o el retiro de electrones
E1 - Piruvato descarboxilasa - coenzima pirofosfato de tiamina TPP - descarboxilación - transfiere el TPP de la enzima al piruvato para que el TPP atraiga a los electrones volviendo el enlace con el CO2 débil, rompíéndose y liberando el CO2 - enzima alostérica - regulable - reacción irreversible
E2 - dihidrolipoil transacetilasa - coenzima ácido lipoico (fuera de la célula: antioxidante (elimina radicales libres y colágeno) - dentro de la célula: oxidación del piruvato facilitando la formación de Acetil-CoA) + CoA. El ácido lipoico se reduce (capta electrones) para oxidar al hidroxietil pirofosfato de tiamina
E3 - dihidrolipoil deshidrogenasa - FAD y NAD+ - le quita los hidrógenos del ácido lipoico
2 son reguladoras: tratan de controlar la forma en la que se activa o se inhibe este complejo.
E4 (piruvato deshidrogenasa cinasa): une o adiciona grupos fosfatos inhibiendo el complejo
E5 (piruvato deshidrogenasa fosfatasa): elimina o quita grupos fosfatos activando el complejo
Estas no utilizan coenzimas
Enzimas: ⬇Energía de activación (para que la reacción se produzca) creando energía de fijación (enlaces débiles entre la enzima y el sustrato)
Transferasas: transferencia de grupos con ayuda de coenzimas
Oxidoreductasas: quitan electrones en forma de hidrógenos con ayuda de coenzimas
Coenzimas: pertenecen al complejo B, favorecen la transferencia de grupos y la eliminación de electrones
REACCIONES COMPLEJO PIRUVATO DESHIDROGENASA
DESCARBOXILACIÓN: la E1 (pirofosfato descarboxilasa) va a transferir el TPP al piruvato aumentando la electronegatividad de la molécula y los electrones se Irán al TPP haciendo que el enlace se rompa y liberando el CO2 - descarboxila al piruvato. Producto = hidroxietil pirofosfato de Tiamina. Se inhibe E1
y 3. FORMACIÓN DE ACETIL - CoA: La E2 (dihidrolipoil transacetilasa) transfiere la coenzima ácido lipoico al hidroxietil pirofosfato de tiamina, reducíéndose captando dos hidrógenos H+ del hidroxietil pirofosfato de tiamina y oxidando al hidroxietil pirofosfato de tiamina, facilitando que dos carbonos (grupo acetilo) se puedan transferir a la CoA, formando Acetil-CoA, esta ya está lista para ir al ciclo de Krebs
4. Y 5. GANANCIA ENERGÉTICA: en la anterior reacción el ácido lipoico (con un alqueno) se quedó con dos H+, entonces la E3 (dihidrolipoil deshidrogenasa) va a tratar de quitarle esos H+ deshidrogenandola con ayuda de un FAD+ (se reduce) que captaran los electrones del ácido lipoico (se oxida) formando FADH2.
FADH2 en la cadena transportadora de electrones genera poca energía, entonces el FADH2 transfiere los electrones a un NAD + para formar NADH para que los lleve a la cadena transportadora de electrones y generar alta cantidad de energía.
Los electrones no pueden pasar directamente al NAD + porque el ácido lipoico es una molécula alquenica, entonces solo FAD+ puede captar sus electrones y no NAD +; si la molécula fuera un alcano entonces sí se podría transferir directo a FAD+.
Ya con el ácido lipoico oxidado, este regresa a la E2, la tiamina regresa a la E1 y ya se puede generar otro ciclo
REGULACIÓN COMPLEJO PIRUVATO DESHIDROGENASA
La E1 es una enzima alostérica y por lo tanto regulable por dos procesos
alosterismo: concentración de moléculas: si aumenta la concentración de NADH (suficiente energía en forma de electrones para formar ATP) o de acetil coA (suficiente energía para generar ciclo de Krebs) se inhibe y se activa de forma contraria, por la baja concentración de NADH, acetil CoA o calcio (a nivel muscular). También se puede inhibir por una alta concentración de ATP y los ácidos grasos porque estos al ser oxidados pueden formar Acetil CoA
Modificación covalente: por adición de grupos fosfatos. La E4 (piruvato deshidrogenasa cinasa) y la E5 son moléculas reguladoras.
E4 (piruvato deshidrogenasa cinasa): une o adiciona grupos fosfatos a la E1 inhibiendo el complejo
E5 (piruvato deshidrogenasa fosfatasa): elimina o quita grupos fosfatos a la E1 activando el complejo
Estas no utilizan coenzimas
Indirectamente la insulina también puede hacer este proceso porque permite que ingrese glucosa cuando hay mucha glucosa y poca energía se mantiene activado; cuando hay mucha insulina y alta energía el complejo se inhibe.
ACIDOSIS LÁCTICA CONGÉNITA - SÍNDROME DE LEIGH - ENCEFALOPATÍA
Estas tres patologías tienen algo en común ya que no permiten que se lleve a cabo el complejo piruvato deshidrogenasa.
En la acidosis láctica no hay una molécula que activa el CPD
En el síndrome de Leigh no se activa la E1
En la encefalopatía no hay TPP adecuada para llevar a cabo el proceso
El piruvato se empieza a acumular, por lo que la lactato deshidrogenasa empieza a formar y acumular ácido láctico que empezara a viajar en la circulación llegando al cerebro, provocando una acidosis metabólica y generando problemas como retraso mental al desnaturalizar proteínas del cerebro, llegando a tener problemas cerebrales, psicomotor o neurológico.
CICLO DE KREBS
o ciclo del ácido cítrico
Hans Adolf Krebs y Fritz Lipman identificaron el complejo aeróbico o ciclo de Krebs, proponiendo que para la degradación compleja de Acetil CoA a CO2 se generan 8 reacciones, cuya última reacción dará un producto que servirá como alimento para generar otro ciclo de krebs.
También se le llama ciclo del ácido cítrico porque el ácido cítrico es el primer producto del ciclo de krebs
También se le llama de los ácidos tricarboxílicos porque los dos primeros productos del ciclo son 3 ácidos carboxílicos.
El ciclo de krebs convergen todos los procesos oxidativos de todas las moléculas que existen. Por ejemplo:
Carbohidrato se degrada llega como acetil coA al ciclo de krebs
Lípido se degrada llega como acetil coA al ciclo de krebs
A.A. Y proteínas se degradan y llegan como acetil coA al ciclo de krebs
Etanol puede llegar a ingresar al ciclo de krebs pero es más difícil que se degrade y solo en bajas concentraciones
Cuerpo cetónicos también convergen al ciclo de krebs
Todo con el objetivo de tratar de oxidar las moléculas obteniendo CO2, aportar moléculas energéticas (electrones); además algunos intermediarios del ciclo de krebs van a servir para la síntesis de otras moléculas, como el citrato sirve para la síntesis de lípidos, el succinil coA sirve para la síntesis de grupos hemo, la alfacetoglutarato sirve para síntesis de aminoácidos, el oxalacetato sirve para la síntesis de moléculas de glucosa. Esto quiere decir que el ciclo de Krebs es una vía anfibólica debido a que provee procesos metabólicos ya anabólicos.
El ciclo de Krebs se divide en tres partes:
OXIDAR EL ACETIL CoA: eliminando sus dos carbonos en forma de CO2
OBTENCIÓN DE ENERGÍA: obteniendo nucleótidos de alta energía y nucleótidos reducidos para atraer electrones y llevarlos a la cadena transportadora de electrones
REGENERAR EL OXALACETATO: para iniciar otro ciclo
REACCIONES
FORMACIÓN DE ÁCIDO CÍTRICO
Tenemos al producto final del ciclo de Krebs anterior Oxalacetato y un Acetil-CoA (proveniente del complejo piruvato deshidrogenasa).
Proceso de condensación aldólica, porque vamos a unir un carbono carbónílico del acetil-CoA con OH del ácido carboxílico del oxalacetato con ayuda de la enzima citrato sintasa (síntesis de un citrato).
Citrato sintasa: enzima de conformación inducible porque solo tiene 1 sitio activo, cuando el oxalacetato se une a este sitio activo, induce a la enzima a que genere un segundo sitio activo para la acetil Co-A. Reacción irreversible - ΔG negativo - libera alta cantidad de energía - exergónica - primera enzima reguladora del ciclo de krebs.
En este proceso participan dos aminoácidos importantes, la Histidina y la ácido aspártico
El ácido aspártico trata de quitar un hidrógeno a la acetil Co-A para que el oxalacetato se pueda unir a la acetil Co-A cuando la molécula se intenta reacomodar para formar citrato, una molécula de agua favorece la ruptura del acetil Co-A de la estructura y esta queda libre, para servir a la otra molécula que viene del complejo piruvato deshidrogenasa; y ahora si se forma el citrato (ácido tricarboxílico - molécula simétrica, sin centros quirales - el ciclo de krebs no puede degradar moléculas simétricas)
FORMACIÓN DE ISOCITRATO - ISOMERIZACIÓN
Como el ciclo de Krebs no puede degradar al citrato porque es una molécula simétrica, entonces debemos isomerizarla para que sea una molécula asimétrica, con ayuda de la enzima Aconitasa que generara un proceso de deshidratación y volverla a hidratar pero en un carbono diferente y así hacerla asimétrica.
La aconitasa es una ferro sulfo proteína: el hierro es el que permitirá el proceso de deshidratación e hidratación.
Entonces, la aconitasa por medio del hierro permite eliminar una molécula de agua formando un inter momentáneo llamado cis-aconitato para luego volver a hidratarla pero colocando los elementos en distinta posición para que sea una molécula asimétrica llamada ISOCITRATO
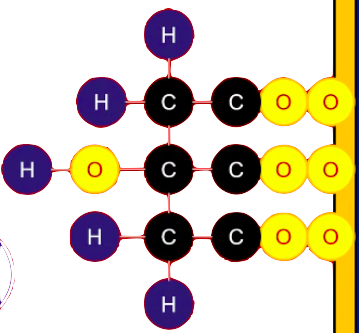
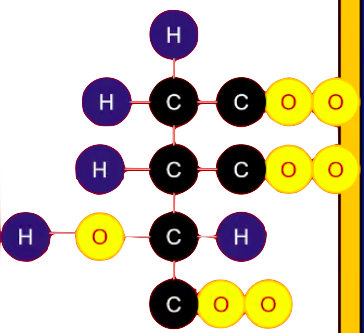
Reacción reversible - ΔG positiva - isostérica
Cuando se consume alta cantidad de flúor (fluorosis por alto consumo de flúor o fluoracetato de las plantas) el flúor puede formar un fluorocitrato que puede detener el ciclo de krebs, ya que se une a la enzima aconitasa y ya no permite que termine el ciclo de krebs.
En la primera y segunda reacción solo preparamos la molécula para ser degradada.
OXIDACIÓN DEL ISOCITRATO - DESCARBOXILACIÓN OXIDATIVA
La enzima Isocitrato deshidrogenasa necesita una coenzima (NAD +) para captar / quitar los electrones hidrógenos del alcohol secundario del isocitrato y convertirlo en una cetona. También puede usar un cofactor (metal inorgánico - magnesio o manganeso) en este caso magnesio que ayudará a que se elimine los electrones.
Entonces el isocitrato deshidrogenasa transfiere un magnesio al isocitrato (se oxida), se forma un oxalsuccinato, se libera un H que se unirá al NAD+ (primer nucleótido reducido) y se debilita el enlace (por la uníón del Magnesio) que une un CO2 y este se libera (primer CO2 liberado); la molécula se estabiliza y el magnesio regresa a la enzima dejando como producto alfa ceto glutarato.
Reacción irreversible - ΔG negativo - alostérica - libera alta cantidad de energía - exergónica - segunda enzima reguladora del ciclo de krebs.
El producto alfa cetoglutarato puede salirse del ciclo de krebs y favorecer la síntesis de aminoácidos (Glu(glutamina, prolina y arginina)) y purinas y alimentar un proceso de transaminación
OXIDACIÓN DE ALFA CETOGLUTARATO - DESCARBOXILACIÓN OXIDATIVA
Aquí usamos un complejo enzimático llamado alfa cetoglutarato deshidrogenasa con E1 (liberación del segundo CO2 - acetil CoA del complejo piruvato deshidrogenasa se termina de degradar), E2 (ácido lipoico quita 2 hidrógenos, forma succinil coA con enlace de alta energía) y E3 (FADH2 y NADH)
NO ES REGULABLE
YA ELIMINAMOS LOS CO2 - TOCA REGENERAR EL OXALACETATO
CONVERSIÓN DE SUCCINIL Co-A EN SUCCINATO - fosforilación a nivel de sustrato
Tenemos a la succinil coA con un enlace de alta energía, necesitamos romper este enlace con ayuda de la enzima succinil CoA sintetasa y sustituirlo con un grupo fosfato que también tendrá un enlace de alta energía, el cual se romperá con la ayuda de la succinil CoA sintetasa, lo sustituye con un OH (formando succinato) y lo traslada a un GDP para formar un GTP luego este GTP trasladará el grupo fosfato a un ADP para formar ATP por medio de la enzima nucleósido difosfato cinasa.
La molécula de succinato puede favorecer la síntesis de propionil CoA (síntesis de nuevas moléculas de glucosa), metionina y ácidos grasos
OXIDACIÓN DEL SUCCINATO
La succinato deshidrogenasa es una enzima integral (proteína que se encuentra pegada en la membrana interna de la mitocondria - unida al complejo II de la cadena transportadora de electrones - asociada porque quita electrones); ayudada por una coenzima FAD+
PREGUNTA DE EXAMEN: ¿Cuál es la única enzima integral del ciclo de krebs?¿Cual es la única enzima unida a la membrana interna de la mitocondria del ciclo de krebs? - succinato deshidrogenasa
Entonces tenemos al succinato le quitamos (se oxida y se forma un alqueno llamado fumarato) un par de electrones en forma de H (con ayuda de la succinato deshidrogenasa) y se los damos al FAD+ (se reduce) para conseguir un FADH2 y éste los transfiere a la cadena transportadora de electrones
HIDRATACIÓN DEL FUMARATO Y PRODUCCIÓN DE MALATO
La fumarasa parte una molécula de H2O en dos formando un grupo OH y un H y que puedan ingresar a la molécula de fumarato para cambiar su configuración producir malato
El malato puede ayudar en la formación de aminoácidos, participa en el ciclo de la urea y favorece la síntesis de purinas
OXIDACIÓN DEL MALATO A OXALACETATO
La malato deshidrogenasa con ayuda del NAD+(se reduce) le quita un H al malato (se oxida) y nos queda un oxalacetato que se podrá volver a usar en otro ciclo de krebs
El oxalacetato puede llegar a formar un fosfoenolpiruvato PEP para la gluconeogénesis
El malato y el oxalacetato es el mecanismo que sirve para trasladar electrones del citoplasma a la matriz mitocondrial
PRODUCTOS
ENTRA | |
Acetil Co-A | del complejo piruvato deshidrogenasa |
(3) NAD+ |
|
FAD+ |
|
GDP |
|
SALE | |
(2) CO2 |
|
GTP |
|
(3) NADH |
|
FADH |
|
Regulación del ciclo de krebs
citrato sintasa (1): alta concentración de NADH, ATP o citrato inhibe la citrato sintasa para que no haya producción de citrato. La succinil co-A también la inhibe (homotrópico). La activa el ATP. Es reguladora porque libera co-A
isocitrato deshidrogenasa (3) (formación de NAD y CO2). Inhibe la alta concentración de ATP. La activa el ATP y la alta concentración de calcio. Regulador por libera CO2
complejo alfa cetoglutarato deshidrogenasa (4): inhibe el NADH y la succinil co-A. La activa el calcio. Regulador porque libera CO2
El ciclo de Krebs está conformado por dos tipos de reacciones importantes:
Reacciones catapleróticas: reacciones que sintetizan moléculas fuera del ciclo de krebs. Cuando salen para producirlas, se detiene el ciclo de krebs
citrato sintetiza ácidos grasos y colesterol
alfacetoglutarato sintetiza ácido glutámico (que sintetizan purinas y aminoácidos)
succinil coA sintetiza grupos hemo
Reacciones anapleróticas: reacciones que alimentan al ciclo de krebs con oxalacetato como intermediario para que cuando se den las reacciones catapleroticas el ciclo no se detenga
TRANSPORTE DE ELECTRONES Y FOSFORILACIÓN OXIDATIVA
CADENA TRANSPORTADORA DE ELECTRONES
NADH, NADPH, Co-A, oxalacetato, iones no pueden atravesar la membrana interna de la mitocondria, por lo que se necesitan un tipo de puertas para que entren los electrones que se formaron en la glucólisis, llamadas lanzaderas (complejos proteicos que van a facilitar el transporte de ciertas moléculas de alta energía que no pueden atravesar la membrana interna de la mitocondria)
Lanzadera glicerol-3-fosfato:
Tenemos un NADH2 y un dihidroxiacetona fosfato. El dihidroxiacetona fosfato capta (se reduce y forma gliceraldehido 3 fosfato) los electrones del NADH (se oxida).
El gliceraldehído 3 fosfato tiene la capacidad de donar los electores al FAD+ de una enzima succinato deshidrogenasa de la membrana, atraviesa la membrana y se forma FADH2. Pasa a la cadena transportadora de electrones
Esta es una reacción rápida, pero menos eficiente ya que necesita mucha energía.
Genera gasto energético.
En cerebro y músculo esquelético
Lanzadera malato-aspartato
tenemos un NADH2 y un oxalacetato en el espacio intermembrana. El oxalacetato capta (se reduce y forma malato) los dos electrones del NADH2 (se oxida) con ayuda de la malato deshidrogenasa.
El malato tiene la capacidada de atravesar la membrana interna por el transportador malato alfa cetoglutarato, ingresando a la matriz mitocondrial, aquí el malato le dona (se oxida y forma oxalacetato) al NAD (se reduce y forma NADH2) dentro de la matriz mitocondrial los electrones que llevaba y se va a la cadena transportadora de electrones.
El oxalacetato que se formó, para regresar al espacio intermembrana sufrirá una transaminación captará un NH2 y se volverá ácido glutámico, así ya puede pasar al espacio intermembrana.
Aquí va a perder ese grupo NH2 y se vuelve oxalacetato
No genera gasto energético
En hígado, riñón y músculo cardiaco
CADENA TRANSPORTADORA DE ELECTRONES
En la membrana interna de la mitocondria existe una serie de complejos proteicos (proteínas integrales, no se pueden mover), con distintas funciones y direcciones. En medio de ellas se encuentran algunas moléculas que sí se pueden mover, entre ellas: la ubiquinona (proteína) y la otra es citocromo c que facilitarán la movilización de electrones.
Dentro de los complejos existen moléculas llamadas acarreadores que servirán para ayudar a entrar y salir del complejo. Estos acarreadores pueden ser nucleótidos o ferro sulfo proteínas o citocromo.
complejo 1 - NADH ubiquinona reductasa : tiene la facilidad de recibir electrones que vengan del NADH y transferirlos por toda la cadena. Usa como acarreador a la ferro sulfo proteína y al mononucleótido de flavina.
Entonces en la entrada tenemos un mononucleótido de flavina (se reduce), el NADH2 le da los hidrógenos (se oxida) y luego esta se los da (se reduce) a varios ferrosulfoproteina (se oxida). Y así se va oxidando y reduciendo en cadena hasta llegar a la salida. En la salida estará la Ubiquinona para recibir los electrones y se traslada al complejo 3
En este proceso por cada electrón que se traslado se liberan dos protones en forma de H+ en el espacio intermembrana
complejo II - : tiene la facilidad de recibir electrones que vengan del FADH2.
FADH le pasa los electrones al succinato deshidrogenasa, este se lo pasa a FAD y este se lo pasa al complejo II, dentro del complejo hay otros FAD que se los irán pasando hasta la puerta de salida, hasta llegar a la ubiquinona y se sigue el mismo proceso, excepto que en el complejo II no se liberan protones H+ y que en el complejo III no se utiliza ferro sulfo proteína
complejo III - ubiquinona citocromo c reductasa: efectora: dentro de ella tenemos como acarreador citocromos B y ferro sulfo proteínas. La ubiquinona le dará sus dos electrones al citocromo B y se irán pasando, reduciendo y oxidando hasta llegar a la salida, donde lo está esperando un citocromo C que captará los electrones
En este proceso por cada electrón que se traslado se liberan dos protones en forma de H+ en el espacio intermembrana
complejo IV: efectora: dentro de ella tenemos como acarreador citocromos A+A3 con hierro+4 y cobre dando un proceso de oxidación y no de reducción, por lo que no necesitaremos una molécula que lo espere al final, solo se unirá al oxígeno que se respira y formará molécula de agua, si no hay oxígeno no habrá molécula de agua
De NADH2 SE FORMARAN 10 Moléculas DE Hidrógeno
DE FADH2 SE FORMARÁN 6 Moléculas DE Hidrógeno
complejo V: ATPasa o ATP sintasa: está dividida en dos partes
F0 proteína integral
F1 proteína libre o periférica, esta está dividida en tres segmentos y aparte cada segmento se divide en un área alfa y en un beta, quedando tres segmentos beta
Beta ATP vacío
Beta ADP + Pi:
Beta ATP
Un primer protón permite la captación de una molécula de ADP y de un fosfato inorgánico en el segmento ADP+Pi, en medio del F1 pasa un protón y la F1 gira y el ADP y el Pi crean una chispa que los une para formar ATP llegando al segmento ATP, pasa otro protón, el F1 gira y se va al segmento Beta ATP vacío, el ATP se libera
Esto se llama fuerza protón motriz, porque se hace que cada protón (3) permita la motricidad o movimiento de la fracción F1 de la ATPasa para sintetizar una molécula de ATP
DE NADH2 SE FORMARAN 3.5 ATP
DE FADH2 SE FORMARAN 2 ATP
REGULACIÓN
Inhibidores del complejo 1: tenemos a la rotenona (insecticida de habas) y al amital (barbitúrico o anestésico). No hay problema si se inhibe porque está el complejo 2
Inhibidores del complejo 2: carboxima (insecticidas de fresas) y malonato (aditivo de lociones). No hay problema si se inhibe porque esta el complejo 1
Inhibidores del complejo 3: antimicina (antibiótico para animales). Puede causar la muerte
Inhibidores del complejo 4: cianuro, monóxido de carbono, ácida. Puede causar la muerte
RENDIMIENTO NETO
ETAPA | PRODUCTO | RENDIMIENTO |
Glucólisis | 2 ATP | 2 ATP |
2 NADH | 5 ATP | |
Oxidación del piruvato | 2 NADH | 5 ATP |
Ciclo de Krebs | 2 GTP / ATP | 2 ATP |
6 NADH | 15 ATP | |
2 FADH2 | 3 ATP | |
TOTAL | 30 - 32 ATP | |